
绝大部分欧亚大陆西部与北非居民都属于体质人类学中的高加索人种,在分子人类学中他们与他们的祖先被称作西欧亚人群。他们的体质特征有一些是来自智人祖型的,比如深眼眶大眼睛。在深目这方面白人比非洲黑人更接近智人祖型。他们也有一些体质特征是最近几万年里逐渐形成的,比如白皮肤、直发与高而窄的鼻子。欧亚大陆西部与北非各族群共同的体貌特征来自基因交流。
上古时期的西欧亚居民包括很多支系,如黎凡特农夫、伊朗农夫、西欧狩猎采集者、东欧狩猎采集者、高加索狩猎采集者等,当时生活在北非的Mechta-Afalou人群与中亚西伯利亚等地的北欧亚人群也与他们有较密切的关系。这些人群之间的差异比现代高加索人种不同族群之间的差异要大,在最近几千年里,由于人群迁徙变得更加频繁,现代欧亚大陆西部及北非不同地区之间的遗传距离已经缩小了很多。现代欧洲人是由欧亚大陆西部不同族群融合形成的,他们有一些遗传性状最初是起源于不同地区的。比如蓝眼睛最初来源于西欧狩猎采集者,Villabruna1、切达人等一万多年前的欧洲古人普遍有着蓝色的眼睛。与欧洲人金发有关的rs12821256最早发现于西伯利亚中部距今1.4万年的北欧亚人群成员Afontova Gora3。与西欧亚人群浅肤色有关的rs1426654则起源于西亚。但需要说明的是,这些遗传性状的扩散并非必须伴随大规模人口替换。尽管rs1426654最初起源于西亚,但早在近东的安纳托利亚农夫大规模迁入欧洲以前它就已经开始在欧洲扩散了。欧洲目前发现的最古老具有这个基因的古人是瑞典Motala遗址7800年前的西欧狩猎采集者。[1]Y染色体单倍群的扩散也是如此,尽管单倍群R起源于中亚,但它也很早就传入欧洲了,目前发现的最古老的R1b是意大利1.4万年前的西欧狩猎采集者Villabruna1。rs1426654的起源地不是欧洲,Y染色体单倍群R的起源地也不是欧洲,但西欧狩猎采集者也可以是R,也可以有浅色的皮肤。之所以会发生这种事是因为人们的祖先数量是很惊人的。我们每一个人都有2个父母,4个祖父母,8个曾祖父母。在往上推的祖先数量是16、32、64、128、256、512、1024……,用不了多少代它就会变成一个天文数字。一般来说,人们的祖先主要来自邻近地区,所以同一个村落、同一座城市、同一族群的人,总是共享更多的基因。但是人们的祖先中也总会有外来人口,因此一些基因可以很快的在那些保持着正常基因交流的群体间扩散也是情理之中的事。
这里特别需要说明的是北非土著也是高加索人种的祖先群体之一。一万年前的西北非土著是Mechta-Afalou人。最著名的Mechta-Afalou人群成员是摩洛哥的Taforalt人,他们生活于1.3万至1.5万年前。Taforalt人的Y-DNA单倍群为E1b1b,线粒体则测出了U6a和M1b,都是现代北非常见的单倍群。不过他们的常染与现代北非人差异较明显。Taforalt人常染有大约63.5%与黎凡特的西欧亚人群有关,另外36.5%与撒哈拉以南非洲相关。[2]相比之下,现代北非人的撒哈拉以南非洲成分要少得多。西亚发展出农业以后人口暴增,周边地区都曾受其移民影响。Taforalt人的确没有竞争过来自黎凡特地区的小麦种植者。但在更早以前,来自非洲的人群其实也对西亚产生过影响,西亚北非与欧洲常见的Y染色体单倍群E1b1b通常被认为是来自非洲的。详情见
阿拉伯语所属的亚非语系也很有可能是来自非洲的,可以参考
的文章。这是Taforalt人的复原图,非常高鼻深目。

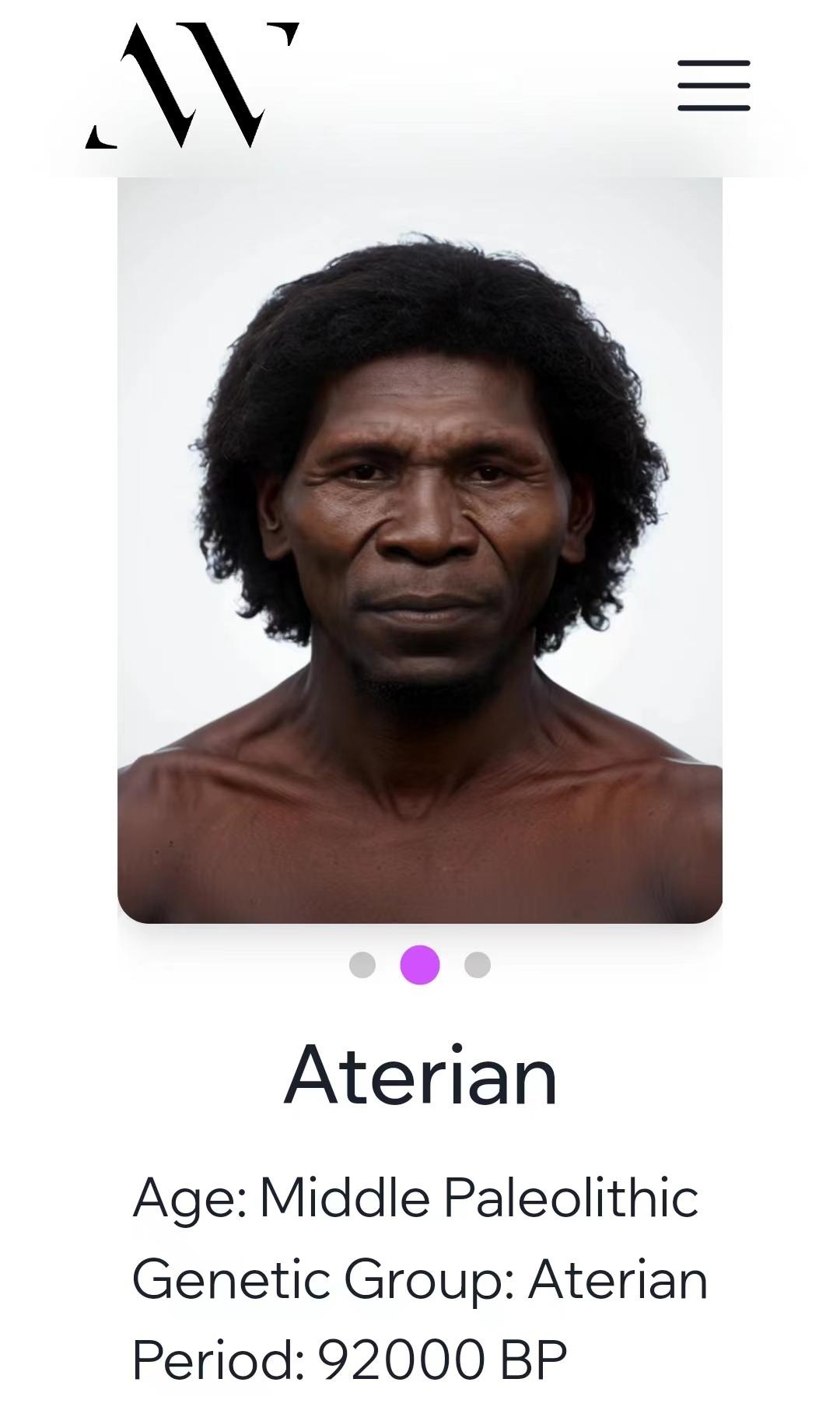
在我们谈论上古时期的晚期智人时,一定要注意这些古人所处的时代。比如这个Arerian人化石是距今9万年的[3],他就不能用来代表北非土著。因为现代人的直系祖先通常被认为是在6.5万年前走出非洲的。9万年前,欧洲还是尼安德特人的地盘,东亚还是丹尼索瓦人的乐土,现代东亚人、欧洲人、澳大利亚原住民与西非黑人的共祖还生活在东非。
目前的DNA与化石证据显示世界上所有现代人都起源于非洲。现代非洲以外人群的祖先来自东非,他们约在6.5万年前走出非洲并逐渐扩散到了整个欧亚大陆与大洋洲,随后由于地理隔离他们被分隔成了几个群体,其中生活在欧亚大陆西部的人群后来形成了高加索人种。欧、亚、非三大洲晚更新世晚期智人化石的形态显示,不同地区的智人在刚走出非洲时具有着相似的外貌,这些共同特征包括低眶、低面、长颅、阔鼻等,古DNA研究则显示他们都拥有较深的肤色。详情参见人类真的起源非洲吗?与智人在走出非洲开始分化以前究竟长什么模样?
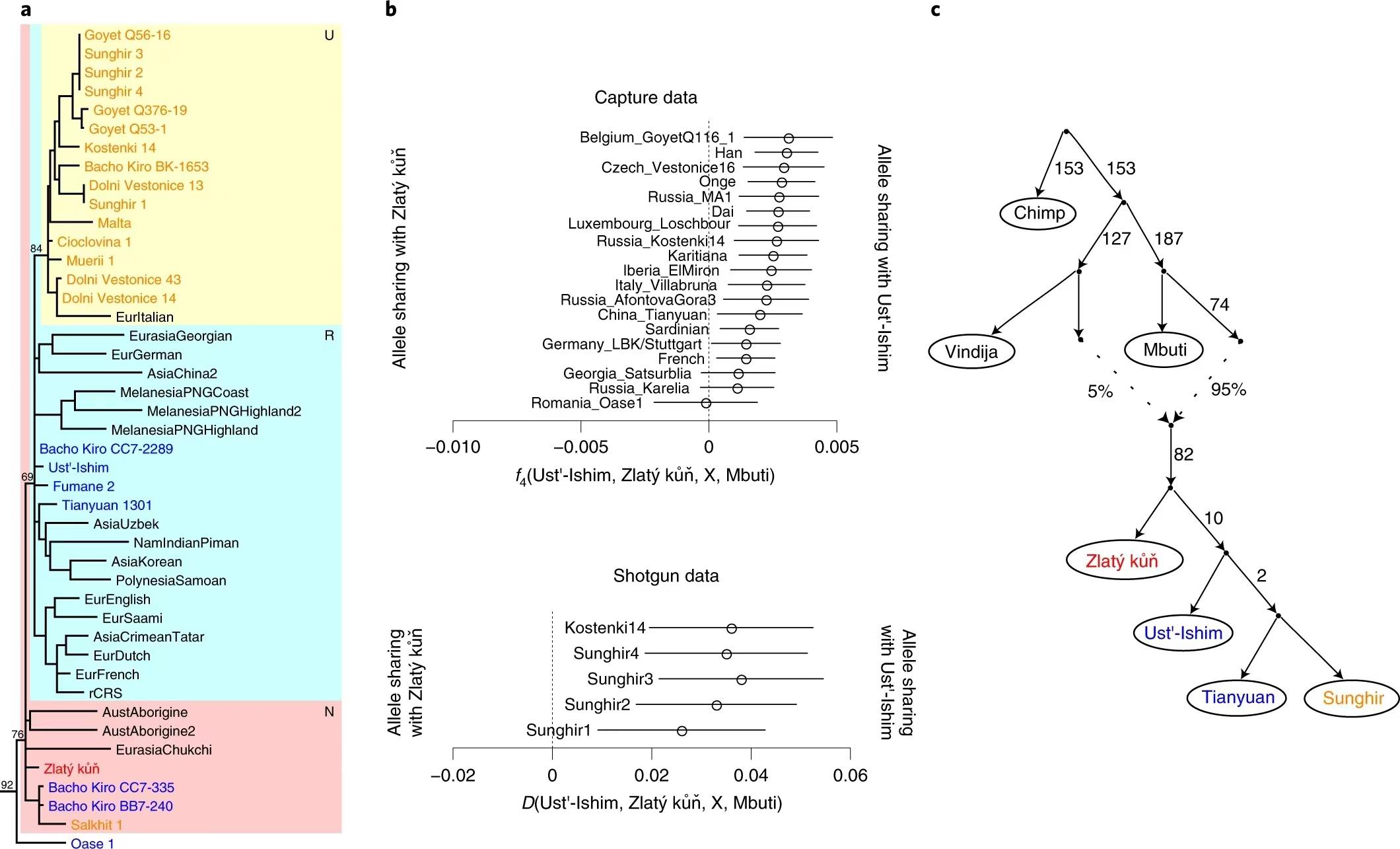
最近7.5万年北半球先后发生过多巴超级火山爆发、坎皮佛莱格瑞超级火山爆发、末次冰盛期、新仙女木事件等多次由气候剧变引起的灾难。许多动物都在这些灾难中灭绝了。我们的祖先非常幸运,他们不但熬过了这些灾难,还在新仙女木事件后发展出了农业。但另一些人群并没有那么幸运,很多人群都在灾难中消失了。没熬过这些灾难的不仅有尼安德特人、丹尼索瓦人,也有一些走出非洲的智人群体。最早走出非洲的智人群体不是我们的祖先,而是Skhul-Qafzeh人,他们十万年前就生活在西亚的黎凡特地区,但是7.5万年前多巴火山爆发后Skhul-Qafzeh人就消失了。6.5万年前走出非洲的这一批人也不是个个都延续了下来,Zlatý kůň与Oase是最早进入欧洲的人群,但是他们在3.9万年前坎皮佛莱格瑞火山爆发后消失了,同时消失的还有与其同时代生活在西伯利亚的Ust-Ishim古人。这张图来自论文《A genome sequence from a modern human skull over 45,000 years old from Zlatý kůň in Czechia》。[4]Sunghir (属于西欧亚人群)与田园洞人(属于东欧亚人群)的分化在他们与Ust-Ishim古人分化之后,他们与Ust-Ishim古人分化又在Zlatý kůň之后。在距今4-5万年前,欧亚大陆上分布着很多支智人族群, Sunghir 所属的西欧亚人群与田园洞人所属的东欧亚人群都是其中一支人群的后代。澳大利亚原住民也是这支人群的后代,他们与东亚人的关系比西欧亚人群更密切。
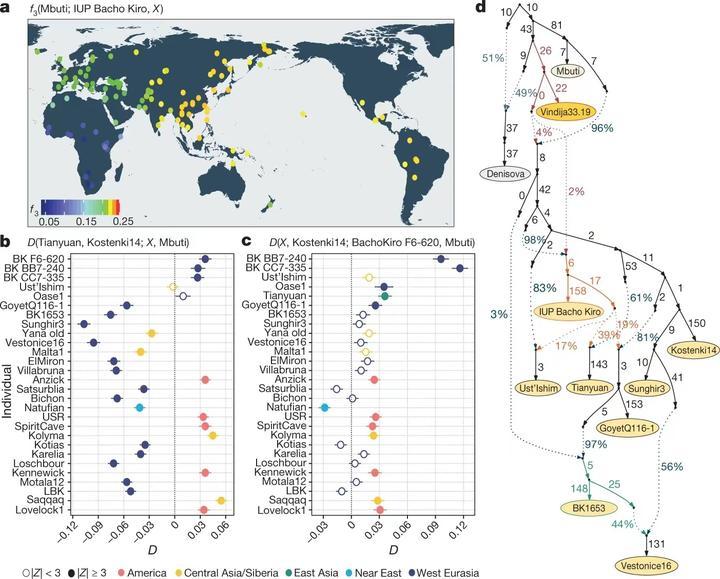
黄白棕种人的祖先在分化以后也与其他人群融合过。西欧亚人群在与东欧亚人群分化以后融合了基底欧亚人(Basal Eurasian)血统,基底欧亚人也属于6.5万年前走出非洲的欧亚人群,但他们很早就和其他人分开了,他们甚至连尼安德特人都没混过。东亚人和澳洲人的祖先在与西欧亚人群分化以后则融合了与Bacho Kiro古人有关的血统。[5]这张图来自《Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry》,其中图a显示的是Bacho Kiro古人与现代各地人群的关系,颜色越红代表关系越密切。相比欧洲人与西亚人,亚洲东部、美洲、大洋洲的族群与Bacho Kiro的关系要近一些,造成这种差异的并不仅是欧洲人与西亚人有基底欧亚人血统,因为四万年前的田园洞古人也比没有混过基底欧亚人血统的欧洲Sunghir古人要接近Bacho Kiro古人。图D显示田园洞人大约有39%的血统接近Bacho Kiro,61%接近Sunghir等西欧亚人群。
虽然东欧亚人群和西欧亚人群在分化后都融合了其他人群的血统,但这种早期的混血可能对他们的外貌影响很有限。因为基底欧亚人与Bacho Kiro古人也是6.5万年前走出非洲的欧亚智人,那个年代的不同人群之间的外貌差异并没有现代人之间的差异大,不同族群的差异是随着时间越来越大的。在不同人群分化以后,相对于智人祖型,所有现代人都发生了变化。其中西欧亚人群与东欧亚人群的变化尤为明显,因为他们生活的地区和非洲的气候环境差异很大。这种改变是逐渐发生的,由一个又一个基因突变累积形成。欧亚大陆西部各地彼此之间有较密切的基因交流,那些有利于适应新环境的衍生等位基因可以很快的在欧亚大陆西部各地扩散。尽管欧洲与西亚、北非的气候并非完全相同,但他们有许多共享的基因,这造就了他们相似的体貌特征。相比之下,欧亚大陆东部与欧亚大陆西部在上古时期存在地理隔离,来往比较有限,因此即便生活在气候环境相似的地区,体貌特征也会出现不小的差异。
东亚人与欧洲人有许多趋同演化形成的遗传性状,这些遗传性状都分别来自不同的基因。最典型的是欧洲人与东亚人都拥有较浅的肤色,然而这两者的浅肤色来源不一样。与欧亚大陆西部的浅肤色有关的rs1426654 衍生等位基因高频分布于欧洲、西亚、北非、南亚、中亚等地,而在东亚少见。与东亚浅肤色有关的rs1800414衍生等位基因则主要分布在欧亚大陆东部,而在欧亚大陆西部很罕见。古DNA研究显示这两个基因突变都是较晚近出现的,欧洲与东亚很多年代较早的晚期智人化石并没有这两个突变。详情见
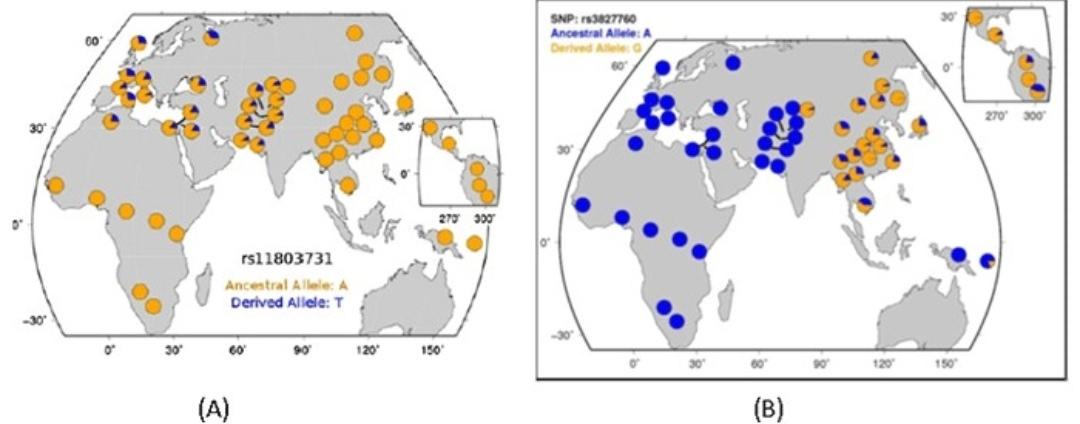
欧洲人与东亚人另一个趋同演化的特征是直发。人类的头发是由多个基因共同控制的,不同族群的头发也不一样。撒哈拉以南非洲黑人与美拉尼西亚人都有非常卷曲的头发。澳大利亚原住民与印度原住民则常见波浪形的头发。东亚人与美洲原住民普遍拥有直发。欧洲人与西亚人的头发介于澳大利亚原住民与东亚人之间,他们中的一部分人有直发,另一部分人有波浪形头发。虽然欧洲人与东亚人都有直发,但这两者的直发来源并不一样。欧亚大陆西部的直发与rs11803731衍生等位基因有关[6],它在欧洲、西亚、北非、南亚、中亚等地以中等频率分布,而在东亚很少见。与东亚人直发有关的rs3827760衍生等位基因正好相反,它高频分布于欧亚大陆东部与美洲各地,而在欧亚大陆西部很少见。欧洲人与东亚人的直发在发质上有很大区别。rs3827760是粗直发基因,它会使头发明显变粗,由于头发直径增加,所以粗直发比卷发或细直发更硬,它也可以使人的发量看上去比实际上更多。
不同种族最显而易见的差异是面部形态,比如高加索人种有非常高的鼻子,东亚人普遍颧骨位置较朝前,人们总是能轻易的分辨出不同种族的面部特征。不过实际上在面部形态方面,不同种族的人之间的共性远远大于他们之间的差异。科学家们在现代人中发现了许多与面部形态有关的基因,这些基因绝大部分都是智人走出非洲以前就具有的,它们同时分布在世界各地的人群中。不管是非洲人、欧洲人还是东亚人,他们之中都有一部分人具有某个基因,另一些人则没有这个基因。只是每个基因在不同族群中的频率存在差异,在一些族群中某个基因占比较高,在另一些族群这个基因占比较低。
下面的表格来自2021年巴基斯坦学者的论文《Selection trends on nasal-associated SNP variants across human populations》,它显示的是在现代人中发现的部分与鼻子形状有关的SNP位点。这里的Afr代表非洲黑人(主要是西非黑人),Amr代表美洲原住民,Ea代表东亚,Eur代表欧洲,Sa代表南亚。这里面有一些位点与鼻子的尺寸有关,它们可以整体的增大或缩小鼻子。还有一些仅和鼻宽有关,可以增加或减少鼻子的宽度。也有一些仅和中庭的长度有关,可以使鼻子变长或变短。还有的与鼻突度有关,可以使鼻子变高或变低。[7]在这些位点中对鼻突度影响较大的是rs2045323。rs2045323衍生等位基因可以增加鼻子的高度,它在非洲黑人中频率仅3%左右,在非洲以外的美洲原住民、东亚人、南亚人中的频率则在20%以上。其他对鼻突度有影响的SNP位点还包括rs11738462、rs1982862等,它们在各族群中的分布也略有差异。人的面部形态是由许多基因共同决定的,单个位点的影响有限。这些已经被发现的基因位点相对所有相关基因位点而言只是很小一部分,还有很多基因还没有被发现,目前单靠已知基因位点是无法推测一个人的外貌的。这张表格的主要意义是让我们了解大部分与面部形态有关的基因都是如何分布的,它们以不同的频率分布在各个族群中,在任何族群中都达不到100%,也不会在任何族群中变成0%。
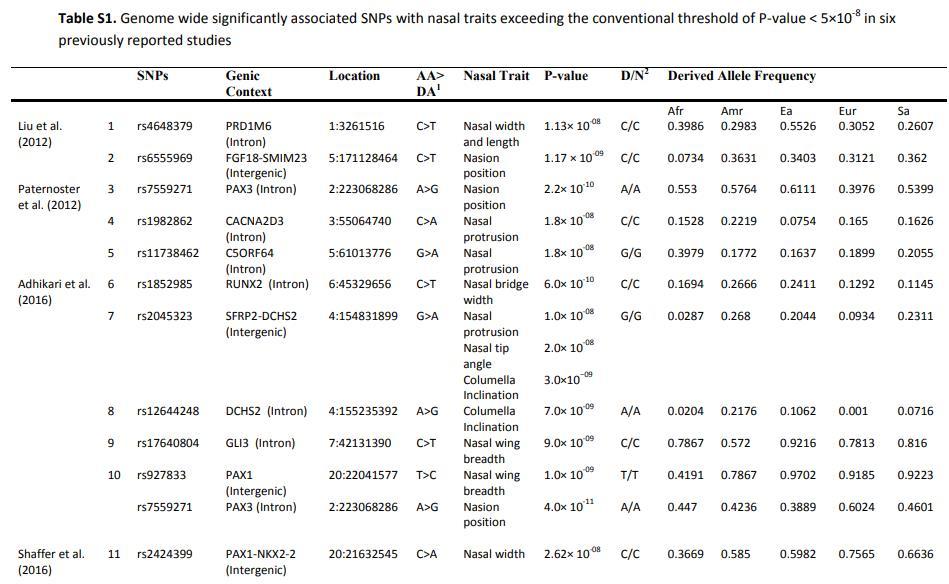
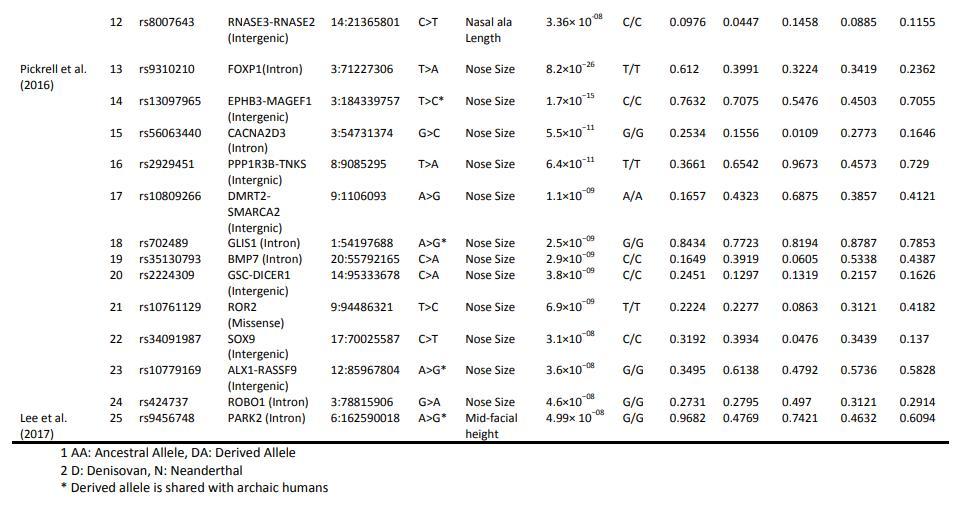
造成欧洲人鼻梁比东亚人更高挺的基因现在还是未知的。2022年,中科院汪思佳、金力领导的团队曾与欧洲科学家合作对欧洲人与东亚人的面部形态差异相关基因进行研究。他们发现rs8068343衍生等位基因在欧洲人与东亚人中的频率有较大区别,它可能会对人的鼻子形状产生影响。不过这个基因并不是欧洲人独有的,它在包括东亚人在内的其他族群中也有分布,而且它的具体影响也有待进一步研究。[8]
前一段时间有媒体报道称现代人的高鼻子基因来自尼安德特人,我看了一下原论文。该论文是由法国国家农业食品与环境研究院与复旦大学专家联合发表的,他们发现来自尼安德特人的基因对一些人的鼻高存在影响。不过体质人类学中的鼻高与人们平常所说的鼻子高度不同,它是指鼻子从鼻根点至鼻下点的距离,即中庭的长度。rs12564392衍生等位基因可以增加中庭的长度,它在美洲原住民及东亚人中频率较高,在欧洲人中频率极低,非洲人没有这个基因。研究者认为这个基因来自尼安德特人,它之所以能在现代人中扩散可能是因为有利于适应寒冷气候。[9]尼安德特人是智人走出非洲时生活在欧亚大陆西部的古人类,他们拥有极长且突出的大鼻子,这不仅与非洲智人有区别,也和同时期的东亚古人不同。现代人的祖先曾经与尼安德特人通婚,以至于非洲以外的现代人普遍具有2%左右的尼安德特人基因。尼安德特人显然有许多可以增加鼻梁高度的基因,也有机会将它们传递给智人,但目前没有证据显示高加索人种有来自尼安德特人的高鼻梁基因。现在已知的与面部形态有关的基因大部分都可以追溯至非洲,仅分布于非洲以外的占比极低。
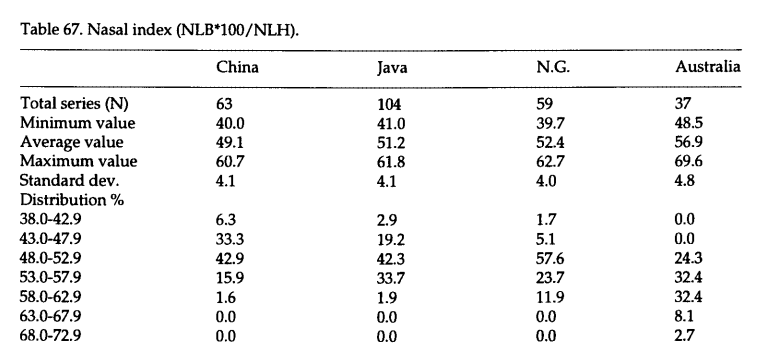
自从十八世纪林奈提出人类亚种的概念以后,人种分类就成了深入人心的概念。一些人类学家为不同种族规定了典型的外貌特征,并认为他们分别起源于不同的地方。对那些不符合XX人种典型形象的族群或个体,则常常被用混血来解释,然而这并非事实,事实是任何族群内部都存在明显的个体差异。在体质人类学书籍中,经常可以看到一些描述种族差异的内容,比如蒙古人种颅型短宽,澳大利亚人种颅型窄长。高加索人种鼻型狭窄,尼格罗人种鼻型宽阔。但这些描述所说的都是平均值,并不是每一个人都符合这些描述。这是美国学者对比尼安德特人与现代人鼻腔差异时统计的不同地区现代人鼻指数数据。鼻指数是根据鼻高与鼻宽计算出来的,它显示的是鼻子的长宽比,鼻子较短或较宽都会导致鼻指数大,较长或较窄则会导致鼻指数小。鼻指数的颅骨测量数据与活体测量数据是不同的,这里显示的是颅骨测量数据。从平均值来看,不同地区的人鼻型差异很明显。非洲黑人普遍有着宽阔的鼻子,他们的鼻指数平均为58左右。北欧人鼻子很狭窄,他们的鼻指数平均为48.2。阿拉斯加因纽特人的鼻宽与北欧人差不多,他们的鼻子长度则明显比北欧人长,所以鼻指数比北欧人更小,平均为45.5。但是除了群体差异之外,个体差异也是显而易见的。北欧人中有一些个体的鼻指数小至39.7,另一些个体的鼻指数则大至59.7。非洲黑人中有一些个体的鼻指数大至69.8,但一些南非人的鼻指数也可以小至47.7。也就是说白人与黑人的鼻指数变异范围存在一定重叠区域。[10]
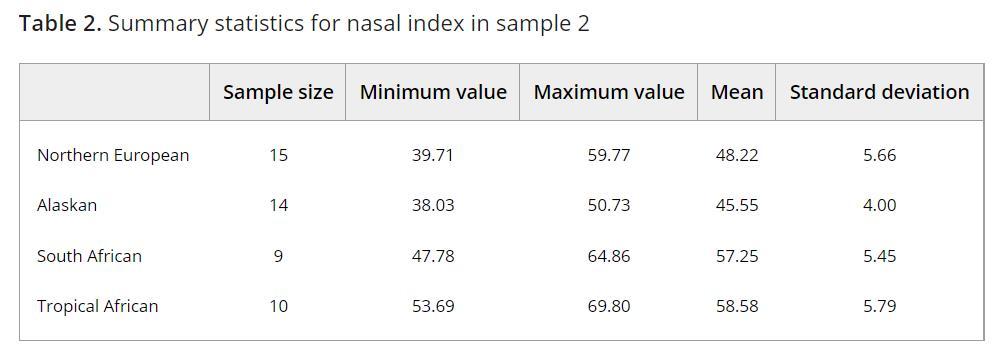
东亚与大洋洲居民的鼻指数变异范围也很广阔。这是《The evolutionary significance of the Wajak skulls》附表中华人、印尼爪哇族、巴布亚人与澳大利亚原住民颅骨的鼻指数统计数据。华人平均鼻指数为49.1。其中鼻指数最小的为40,最大的为60.7。澳大利亚原住民平均鼻指数为56.9。其中鼻指数最大的为69.6,最小的为48.5。文中的另一些附表显示各族群的鼻宽绝对值变异范围也同样很广阔。华人男性鼻高平均为52.3mm,鼻宽为25.6mm。其中鼻宽最大的可以达到30.6mm,最小的仅21.9mm。澳大利亚原住民男性鼻高平均为50.2mm,鼻宽为28.8mm。其中鼻宽最大的可以达到34mm,最小的仅24.2mm。[11]
鼻型短宽的白人与鼻型窄长的黑人是比较少见的,但少见不等于没有。比如芬兰前总理尤哈·西皮莱的鼻型就比卢旺达总统卡加梅短宽的多。尤哈·西皮莱的鼻型比较宽阔并未妨碍人们一眼就能认出他是欧洲白人,这是因为人种不是由单项特征决定的。尤哈·西皮莱虽然鼻型短宽,但鼻子高度也较高。他的深眼眶、正颌、薄嘴唇、直头发、白皮肤、较浅的眸色等等都是典型的欧洲白人特征。白人中也有嘴唇厚的,比如本拉登的嘴唇就相当厚,但他仍然一看就是正宗的西亚白人,这也是同样的道理。

非洲黑人的相貌多样性也比大部分人想象中要高的多,实际上根据一些学者的研究,非洲黑人同族内部的颅骨形态变异范围比其他人更广阔。Lahr曾经将非洲黑人称作(现代人中)最异质的人群。[12]对卢旺达有所了解的人都会知道卢旺达人可以被分为图西族与胡图族,他们在外貌上存在区别,然而实际上胡图族与图西族在种族血统上究竟有多少区别是很难说的。下图是卢旺达人的不同相貌类型。比利时殖民者曾经认为上面三种类型是图西族,下面三种类型是胡图族。并认为图西族与埃塞俄比亚人有关,两者都有白人血统。但是现代分子人类学研究显示图西族父系与胡图族相似,均以E1b1a为主,兼具部分B。至少在父系方面,图西族不但与白人毫无关系,也不接近非洲之角。[13]我没有看到图西族的母系情况,但既然父系和白人毫无关系,估计母系也不会有多少白人血统。
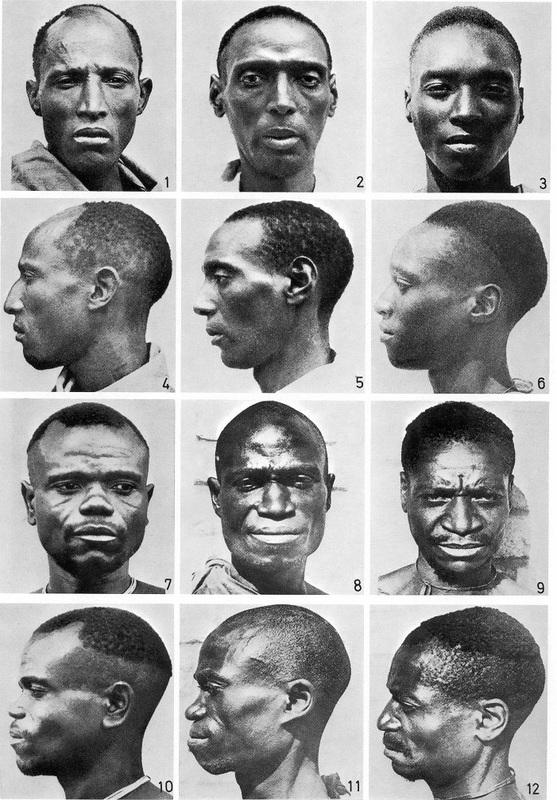
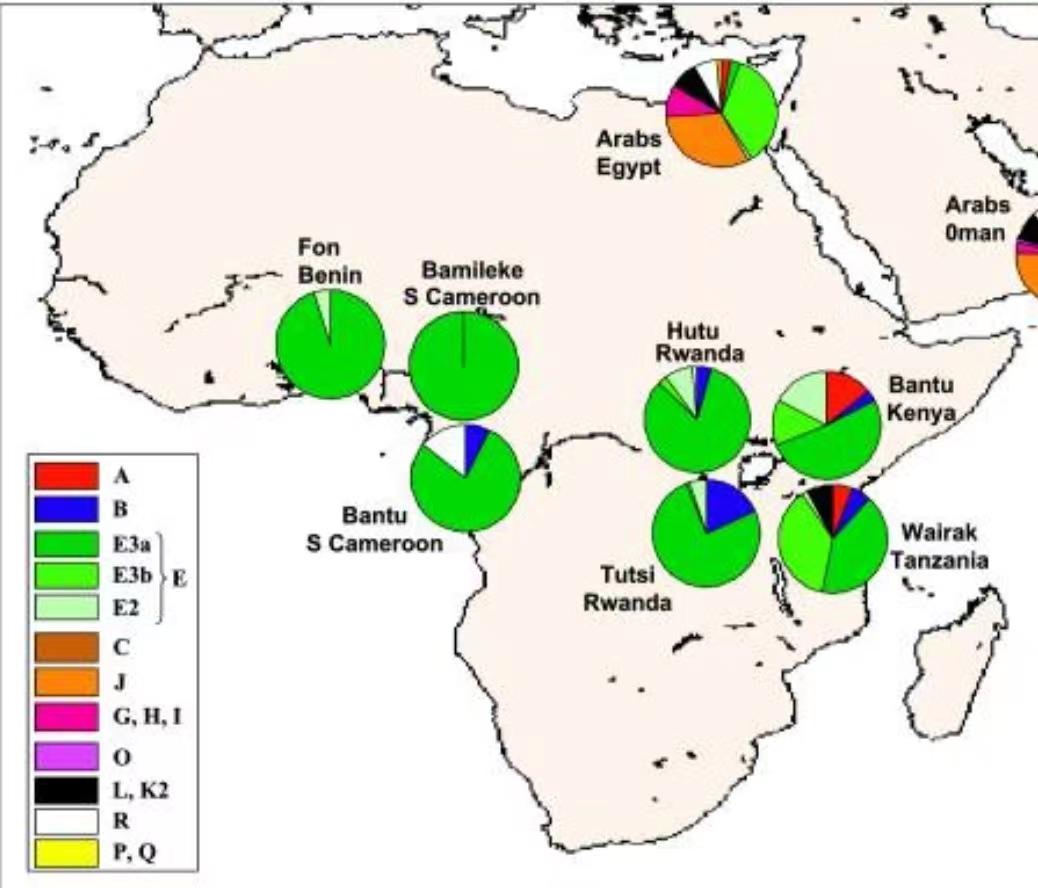
这是不同Y-DNA单倍群在非洲部分族群中的比例。E3a现在叫做E1b1a,是撒哈拉以南非洲的主要Y-DNA单倍群,在尼日尔-刚果语族中高频,它也是卢旺达人中居主导的单倍群。E3b现在叫做E1b1b,是非洲之角的主要Y-DNA单倍群,它在卢旺达人中占比很低。图西族与胡图族的区别是B占比稍高。
除了卢旺达人,其他黑人族群内部也都具有很高的多样性。在所有非洲黑人族群中,非洲之角居民与走出非洲的欧亚人群具有最密切关系。由于与欧亚大陆交流密切,现代非洲之角居民大多属于黑白过渡人种,但这里也有不属于黑白过渡人种的族群。埃塞俄比亚西南部的奥莫山谷生活着世界上最后的原始部落群体之一,这些部落民世代隐居在山谷中,直到七十年代才被外界发现,他们是典型的非洲黑人族群。非洲之角的地形十分多变,这里既有高原,也有低谷,既有森林、草原,也有沙漠,既有气候干燥的地区,也有湿润的地区。其中索马里以热带沙漠居多,埃塞俄比亚则以气候湿润的高原、山地为主。奥莫山谷在东非大裂谷附近的低海拔地区,那里气候炎热,湿度适中。

奥莫山谷部落民相貌很多样,他们中的大部分人和通常印象中的典型黑人一样,窄脸大眼睛,鼻子比较宽塌,嘴唇较厚。但是他们中也有鼻梁较高挺的人,还有少数人具有内眦褶与内双。这是鼻梁较高挺的Hamar族男子。

拥有内眦褶与内双的Hamar族女子。

白人内部也存在不同的相貌类型。这是一种白人常见的相貌,过去的一些体质人类学研究者称之为armenid,亚美尼亚型,最明显的特征就是拥有超高的鼻子。虽然被称作亚美尼亚型,但这种相貌并不是高加索地区的专属,在欧洲、西亚及北非各地都可以看到这种鼻子超高的相貌。这些人也未必具有特别密切的关系,高加索人种中各类增加鼻梁高度的基因频率比其他人高,有时候叠加的高鼻梁基因太多了就会出现鼻梁超高的情况。

这也是一种在欧洲常见的相貌,过去的一些体质人类学研究者称之为East Baltic,东波罗的海型,在白人中属于五官相对小巧的。这种相貌在欧洲(尤其是东欧与北欧)很常见,在西亚比较少。

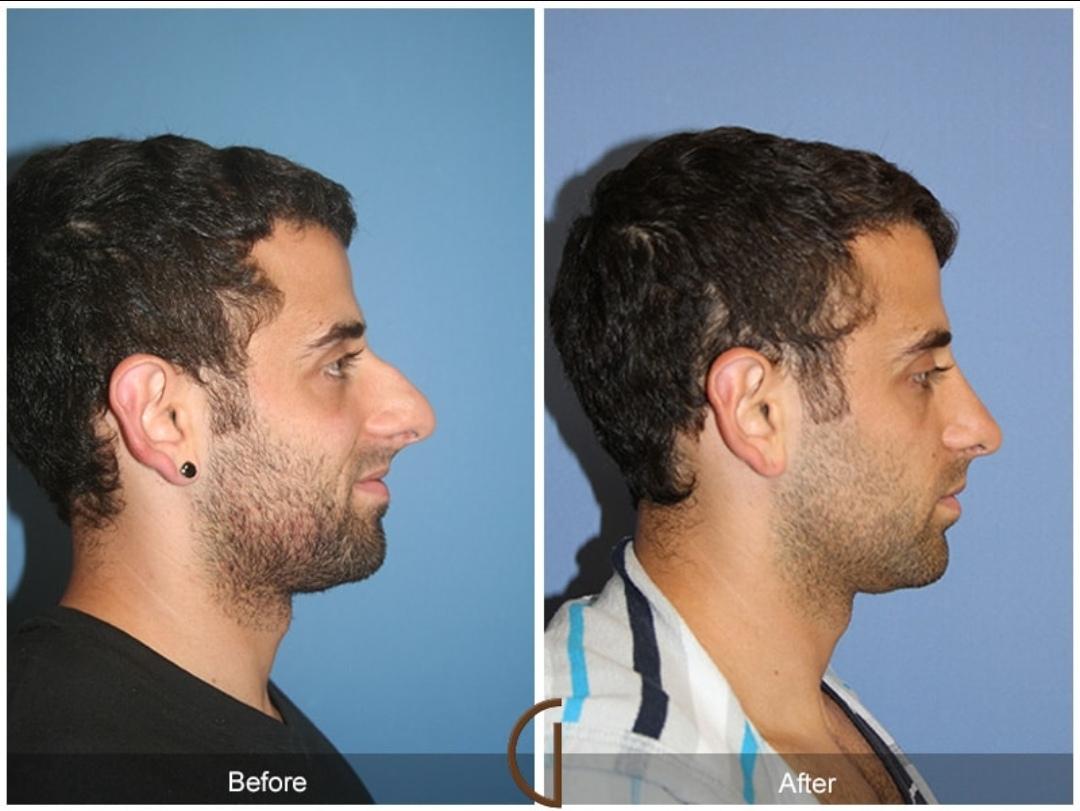
就像东亚流行垫高鼻子的整容手术一样,白人中也流行将鼻子削小的整容手术。很多拥有大鹰钩鼻的白人会特意去将自己的鼻子削成与好莱坞明星同款的希腊鼻。这种手术在伊朗等西亚国家尤为流行,因为西亚人的鼻子平均来说比其他地区的白人更高,他们更容易长出这种超高的鼻子。这是缩鼻手术前后对比。
东亚人内部同样存在个体差异,下图这两种相貌在东亚都有分布。比如我国诺贝尔文学奖得主莫言就比较接近第一个人,另一个华人诺贝尔文学奖得主高行健则比较接近第二个人。华人诺奖得主总共也没几个,在他们中都能同时找到这两种类型的,可见这两者在中国都不算少见,但这两者之间的差异也是很明显的。每个族群的每项颅骨测量数据都会有一个平均值,它对应的是相关基因在不同族群中的不同频率。但放在每个个体身上,每个人的测量数据并不会都像平均值一样,它有一个变异范围。总会有人的数值比平均值更高,也总会有人比平均值更低。
美洲原住民内部也存在明显的个体差异。比如亚马逊雨林原住民既有像左边这样鼻梁比较低的,也有像右边那种鼻梁高挺的,大部分人则介于这两者之间。

在蒙古人种中,美洲原住民的鼻梁高度平均值比亚洲蒙古人种要高一些,这在美国、中美洲与安第斯山脉等地的原住民中尤为明显。他们有时甚至会出现高的过分的鼻子。比如这位名为Long Bull 的苏族酋长有个外号叫大鼻子,这种大鼻子在亚洲蒙古人种中是见不到的。这种相貌能在美国原住民中出现,说明加高加大鼻子的基因在美国原住民中的频率比东亚更高。东亚人与美洲原住民的颅骨测量数据是比较接近的,因此常常撞脸。但这两者的颅骨测量数据也不尽相同,所以彼此也会出现对方所没有的相貌。

值得一提的是,美洲原住民刚抵达美洲时和现在是有区别的,就像当时的东亚人也和现在有区别一样。美洲最早的晚期智人化石被称作Paleo-american。与同时期的东亚古人相似,相比现代东亚人或美洲原住民,这些古美洲人带有更多的智人祖型特征。如颅型更长,鼻型较短宽,眼眶更低矮。[14]不过古DNA研究显示这些古美洲人正是美洲原住民的祖先。[15]这或许意味着,美洲原住民在与东亚人分化以后,两者仍有一些趋同演化,包括颅型向短宽发展、鼻型向窄长发展、眉骨弱化等。这是古美洲人与现代美洲原住民颅骨对比。
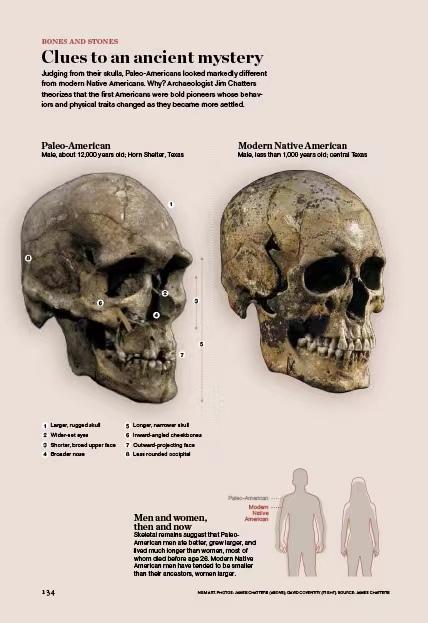
这是古代美洲居民的复原图。其中前七个是Paleo-american,后面都是体质人类学意义上的现代美洲原住民。古美洲人的相貌很多样,既有Naia那种鼻子较低的类型,也有肯尼威克人那样鼻子特别高的类型。

Naia与肯尼威克人的复原像,肯尼威克人长着非常高的鹰钩鼻,Naia正好相反,但他们都是古美洲人。

面对种族差异,人们很容易出现两种倾向。一个是将不同种族视作完全不同的亚种,连偶尔出现与对方相似的相貌都必须是混血。另一个是忽视遗传漂变的可能性,认为不同人群在基因交流很少的情况下也应该完全保持一致,如果不一致就一定是起源于不同的环境。最典型的例子是很多人坚信高加索人种与蒙古人种一定分别起源于不同的气候带,如果一个是起源于温带,另外一个就必须起源于寒带。对于高加索人种与蒙古人种的外貌差异,有两种截然不同的说法,一种说法称高加索人种的鼻梁比蒙古人种更高是因为他们更适应寒冷干燥的环境。另一种说法则认为蒙古人种比高加索人种更适应严寒,高加索人种的高鼻来自对干热沙漠气候的适应。然而这些说法都缺乏证据。因为高加索人种内部的阿拉伯人与北欧人对环境的适应是不同的。同属蒙古人种,东南亚人或中南美洲原住民与因纽特人对环境的适应也是不同的。没有证据表明亚马逊雨林原住民比北欧人还要适应寒冷气候,就像没有证据表明阿拉伯人比因纽特人更适应寒冷气候一样。
2015年美国学者Arslan. Zaidi等人曾经对西非、东亚、北欧、南亚次大陆居民的鼻部进行测量并对各项数值与当地气候的关系进行过研究。他们认为各族群鼻子形状差异的某些方面确实是出于当地对气候的适应,但是否所有方面都完全取决于气候则有待商榷。不同族群的鼻子形状差异可能还受其他力量影响,如性选择,将之全部解释为对气候的适应是对复杂演化史的简单化。[16]这是测量数据,需要注意的是这是活体测量数据,与前面的颅骨测量数据是不同的。
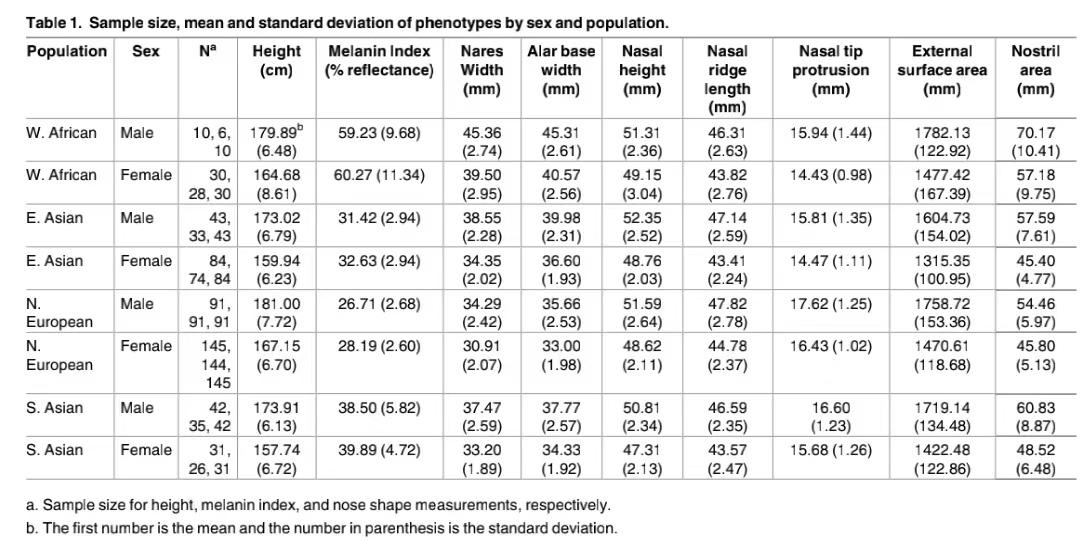
Arslan. Zaidi等人的研究显示,在鼻部的各项测量值中,与气温关系最密切的是鼻孔面积。在所有参与测量的族群中,西非黑人的鼻宽远比其他人宽,鼻孔面积也远大于其他人,男性达到了70平方毫米。白人都有较高窄的鼻型,但是来自不同地区的白人鼻孔面积并不一样。北欧男性的鼻孔面积为54平方毫米,南亚男性的鼻孔面积则为60平方毫米。东亚人的鼻孔面积介于北欧与南亚之间,为57平方毫米。[16]
Arslan. Zaidi等人还对比了不同地区的人的鼻宽与当地湿度之间的关系,因为此前曾有人指出干燥气候会驱使人的鼻子朝高窄型演化。他们认为鼻宽可能与绝对湿度(而不是相对湿度)存在关联,只是这种关联并不像肤色与紫外线的关联那样紧密。他们还指出目前参与研究的族群数量有限,(美国的移民祖籍地以欧洲、西非、东亚、南亚为主,欧美对于种族差异的研究也往往局限于这几个族群。)要得出准确的结论需要更多族群参与。尤其需要检测的是青藏高原、南美安第斯高原以及东非埃塞俄比亚高原居民的情况,因为他们生活的高海拔地区不仅气候与同纬度地区存在差异,还会涉及高原缺氧问题。[16]

非洲大部分地区均属热带,这很容易让人以为这些地方都有相似的气候,都是干热或湿热的。但实际上非洲的气候也很多元,不同地区的气候并不一样。不但东非、西非、南非有区别,东非、西非、南非内部的不同地区也存在差异。西非沿海地区以热带雨林为主,内陆则主要为热带草原。东非的地形与气候更加复杂。那里既有酷热的索马里沙漠和尼罗河上游盆地,也有凉爽的埃塞俄比亚高原。详情见
参考
- ^How Europeans evolved white skin https://www.science.org/content/article/how-europeans-evolved-white-skin
- ^Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations https://www.science.org/doi/10.1126/science.aar8380
- ^Ancestral Whispers https://www.ancestralwhispers.org/reconstructions/aterian
- ^A genome sequence from a modern human skull over 45,000 years old from Zlatý kůň in Czechia https://www.nature.com/articles/s41559-021-01443-x
- ^Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry https://www.nature.com/articles/s41586-021-03335-3
- ^Common variants in the trichohyalin gene are associated with straight hair in Europeans https://europepmc.org/article/pmc/pmc2775823
- ^Selection trends on nasal-associated SNP variants across human populations https://www.sciencedirect.com/science/article/pii/S2214540021000232#t0010
- ^Genetic mechanisms underlying East Asian and European Facial differentiation https://www.researchgate.net/publication/352702464_Genetic_mechanisms_underlying_East_Asian_and_European_Facial_differentiation
- ^Automatic landmarking identifies new loci associated with face morphology and implicates Neanderthal introgression in human nasal shape https://www.researchgate.net/publication/370596197_Automatic_landmarking_identifies_new_loci_associated_with_face_morphology_and_implicates_Neanderthal_introgression_in_human_nasal_shape
- ^The Nasal Complex of Neanderthals: An Entry Portal to their Place in Human Ancestry https://anatomypubs.onlinelibrary.wiley.com/doi/10.1002/ar.23040
- ^The evolutionary significance of the Wajak skulls https://www.researchgate.net/publication/254911892_The_evolutionary_significance_of_the_Wajak_skulls
- ^中国与非洲近代-现代人类某些颅骨特征的对比及其意义——中国与非洲人类头骨特征对比之二 http://www.ivpp.ac.cn/cbw/rlxxb/xbwzxz/201202/t20120221_3443232.html
- ^The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1182266/
- ^Craniometric evidence for Paleoamerican survival in Baja California https://www.researchgate.net/publication/10583538_Craniometric_evidence_for_Paleoamerican_survival_in_Baja_California
- ^Ancient DNA tracks migrations around Americas https://www.science.org/doi/10.1126/science.362.6415.627
- ^abcInvestigating the case of human nose shape and climate adaptation https://pdfs.semanticscholar.org/a69f/b8f6bfa764d301e8eb899bccd9f6fe898db0.pdf?_gl=1*dgsd2k*_ga*MzEwNDAzNjQ2LjE2NjgyMjY1NTc.*_ga_H7P4ZT52H5*MTY4NTMyNjQzMy4yLjAuMTY4NTMyNjQzNi41Ny4wLjA